Les membranes photosynthétiques sont si abondantes (1 m2 de feuilles contient une surface photosynthétique supérieure à celle d’un stade de rugby), que les galactolipides, briques de base des membranes biologiques, sont les lipides polaires les plus abondants sur Terre. Ces molécules polaires sont constituées d’une tête hydrophile et d’une queue hydrophobe constituée de 2 longues chaînes de carbones appelées acides gras (AG). Chaque membrane de la cellule a une composition en glycérolipides qui lui est propre : les chloroplastes des cellules végétales et des algues sont caractérisés par la présence d’une classe de glycérolipides uniques, les galactolipides. Dans une cellule chlorophyllienne la synthèse
de novo des AG est initialement réalisée dans le stroma des chloroplastes (alors que dans les cellules animales, elle a lieu dans le cytoplasme). Il existe deux sites principaux d’assemblage des glycérolipides membranaires (générant directement des membranes) : l’enveloppe des chloroplastes et le réticulum endoplasmique.
Chez les plantes, les galactolipides, synthétisés dans le chloroplaste, peuvent porter 2 AG de 16 ou 18 atomes de carbone. Chez les microalgues et les diatomées, les galactolipides sont riches en AG polyinsaturés à très longues chaînes, dont l’acide eicosapentaénoïque, appelé aussi 20:5 car constitué de 20 carbones et de 5 doubles liaisons. Si le chloroplaste peut en théorie synthétiser des AG de 16 ou 18 atomes de carbone, il ne peut pas en produire de plus longs ; la biosynthèse du 20:5 est cytoplasmique. Plusieurs enzymes de synthèse de 20:5 sont connues : des élongases qui ajoutent 2 carbones à un AG, et des désaturases qui ajoutent les doubles liaisons. Il était admis que le substrat initial des enzymes cytoplasmiques était le 18:2 qui subissait une élongation et trois désaturations pour arriver à du 20:5. Or, chez les microalgues telles que
Nannochloropsis, des élongases sont capables d’accepter du 16:0 comme substrat initial. A quoi sert un système d’élongation permettant de passer de 16 à 18 atomes de carbones dans le cytoplasme ? Quel est le rôle de ces élongases ?
Pour répondre à ces questions, nous nous sommes intéressé à la famille de 6 élongases cytoplasmiques de Nannochloropsis (Δ0-ELO1 à 6) catalysant la synthèse de 18:0 à partir de 16:0. Par une étude de génétique fonctionnelle, nous avons montré que Δ0-ELO1, la plus exprimée d’entre elles, était le point d’entrée des AG vers un système multi-enzymatique qui permet d’aboutir au 20:5, dirigé ensuite vers le chloroplaste où il est spécifiquement utilisé pour produire un galactolipide chloroplastique. De ces données, nous avons obtenu un mutant n’exprimant pas Δ0-ELO1 (
Figure) et dans lequel ils observent la baisse de la teneur du galactolipide en question, et par effet d’aiguillage, une production d’huile obtenue à partir de l’accumulation de 16:0.
Ces travaux permettent non seulement de répondre à des questions fondamentales concernant la synthèse très complexe des AG chez les microalgues, mais mettent également en évidence un moyen de provoquer génétiquement une accumulation d’huile chez
Nannochloropsis. Ils font l’objet d’un dépôt de brevet avec la société Fermentalg.
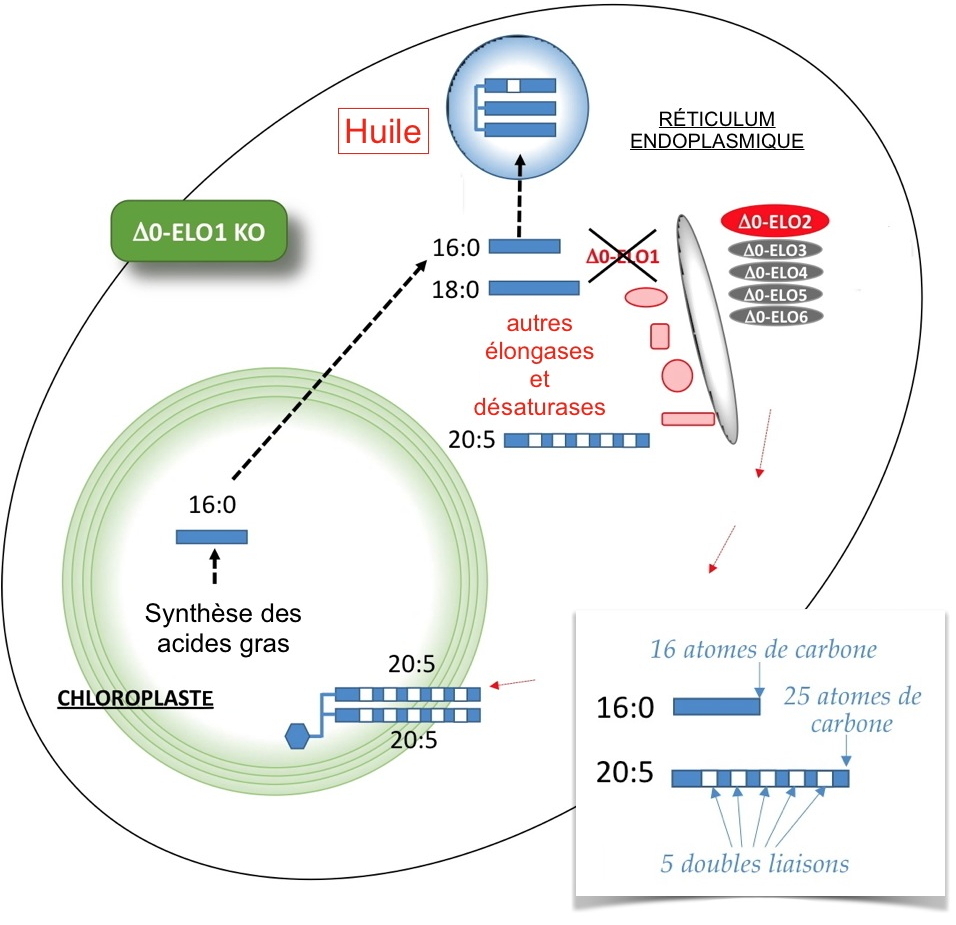
Le 16:0 chloroplastique est normalement exporté dans le cytosol où il est pris en charge par des élongases et des désaturases afin de donner du 20:5 qui est ensuite redirigé dans le chloroplaste.
Dans ce mutant ne produisant pas Δ0-ELO1, du 16:0 s’accumule et est dans ce cas dirigé vers la production d’huile, ou triacylglycérol contenant 3 AG.