Dans une cellule eucaryote, chaque membrane a une composition lipidique (ou profil lipidique) spécifique.
La question générale posée par l’équipe est de déchiffrer les processus moléculaires responsables de l’établissement et de la conservation des profils glycérolipidiques dans une membrane et dans une gouttelette d’huile, dans des organismes eucaryotes contenant des plastes (plantes et algues). Les glycérolipides sont faits à partir de l’assemblage d’acide gras (AG), de glycérol et de tête polaires. Moins d’une douzaine de classes de glycérolipides sont suffisantes pour composer la plupart des membranes biologiques. Cela inclut les phospholipides tels que la phosphatidylcholine, la phosphatidyléthanolamine qui sont les constituants principaux des membranes extraplastidiales, et les lipides non phosphorés tels que les galactolipides (monogalactosyldiacylglycérol et digalactosyldiacylglycérol) qui composent la majorité des membranes photosynthétiques (Figure 1). Chez les algues, il existe un troisième type de lipide membranaire non phosphoré, appelé bétaïne lipide, qui serait aussi localisé dans les membranes extraplastidiales.
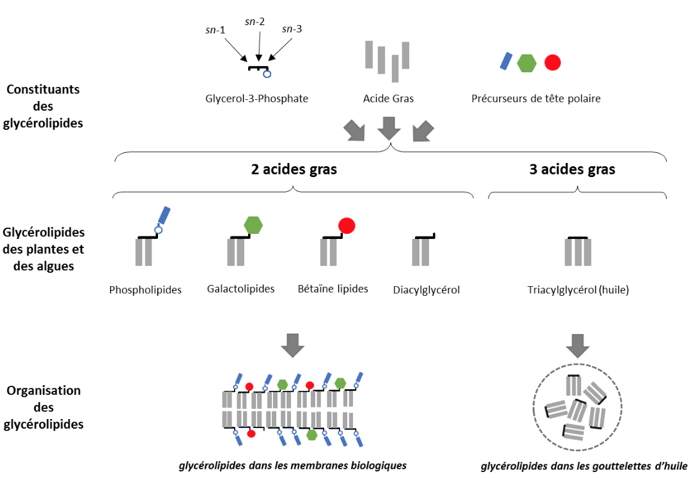 Figure 1 : Les glycérolipides, constituants des membranes biologiques et des huiles. Les glycérolipides sont un groupe particulier de lipides qui contiennent un squelette glycérol sur lequel de 1 à 3 acides gras peuvent être estérifiés. Les glycérolipides qui contiennent deux acides gras peuvent avoir une tête polaire et sont capables de s’organiser en bicouche, formant ainsi la base des membranes biologiques. Les glycérolipides contenant trois acides gras sont appelés triacylglycérol, ou huile, et s’accumule sous la forme de gouttelettes à l’intérieur de la cellule.
Figure 1 : Les glycérolipides, constituants des membranes biologiques et des huiles. Les glycérolipides sont un groupe particulier de lipides qui contiennent un squelette glycérol sur lequel de 1 à 3 acides gras peuvent être estérifiés. Les glycérolipides qui contiennent deux acides gras peuvent avoir une tête polaire et sont capables de s’organiser en bicouche, formant ainsi la base des membranes biologiques. Les glycérolipides contenant trois acides gras sont appelés triacylglycérol, ou huile, et s’accumule sous la forme de gouttelettes à l’intérieur de la cellule. Chez les plantes, la biosynthèse des glycérolipides répond schématiquement à 4 règles :
- La biosynthèse des acides gras a lieu dans le stroma des chloroplastes ;
- L’assemblage des acides gras et du glycérol pour former les phospholipides se déroule dans le réticulum endoplasmique ;
- La synthèse des galactolipides, sulfolipides et d’une partie du phosphatidylglycérol se déroule dans l’enveloppe du chloroplaste ;
- Les glycérolipides membranaires peuvent être convertis en triacylglycérol, une réserve de lipides stockée dans des gouttelettes (Figure 1). Ainsi, la composition lipidique des membranes dépend de 3 phénomènes essentiels : (1) la néosynthèse des glycérolipides ; (2) la conversion entre les glycérolipides, par échange de tête polaire ou d’acides gras et enfin (3) le transport ou trafic de glycérolipides à l’intérieur de la cellule.
Dans une cellule eucaryote, chacun des compartiments membranaires (membrane plasmique, membranes du réticulum endoplasmique, du Golgi, de la vacuole, des mitochondries, des plastes, etc) présente une composition lipidique qui lui est spécifique et qui est très stable. En terme physiologique, cet état d’équilibre nécessite une série complexe de processus que nous appelons « homéostasie des lipides membranaires », et qui est la trame de fond de l’intégralité des recherches conduites par l’équipe.
Des modifications de la composition des membranes peuvent intervenir en réponse à des variations de conditions environnementales. Ainsi, chez un organisme photosynthétique dans un environnement carencé en phosphate, on observe un appauvrissement de certaines membranes en phospholipides, contrebalancé par leur enrichissement en galactolipides. L'ensemble représente à l'échelle cellulaire une économie vitale en phosphate. Un transfert de galactolipides se produit des membranes des chloroplastes vers d'autres membranes habituellement dépourvues de galactolipides telles que les membranes mitochondriales. Les processus de couplage entre voies de synthèses de phospholipides (dans le réticulum endoplasmique) et de galactolipides (dans l'enveloppe des chloroplastes), les mécanismes régulateurs et les machineries de transferts de lipides sont méconnus.
L’équipe a une solide connaissance dans le domaine des glycérolipides végétaux.
Chez les plantes les enzymes de synthèse et de modification des lipides sont bien connus. En ce qui concerne les microalgues le socle de connaissances est beaucoup plus faible et nécessite une investigation de toutes les bases du métabolisme lipidique. Les microalgues regroupent des organismes issus de l’endosymbiose primaire (Archaeplastida), à l’instar des plantes, mais également de l’endosymbiose secondaire (Chromista).
Nos modèles d’endosymbiose primaire sont la plante
Arabidopsis thaliana,
Ostreococcus tauri ainsi que les Chlorophyta se développant aux très basses températures (psychrophiles), telles que
Sanguina nivaloides. A l’instar des autres eucaryotes, le trafic lipidique et les systèmes de tri des plantes sont à peine compris, une lacune de connaissance que nous nous attelons à combler grâce au modèle
A. thaliana.
La picoalgue
O. tauri est un système végétal minimal et un organisme marin à la croisée évolutive des des deux principaux règnes de microalgues (Archaeplastida, Chromista). C’est un modèle idéal pour l’étude des réseaux de gènes sous-tendant la reprogrammation du métabolisme lipidique spécifique des microalgue marines en réponse aux stress abiotiques.
Nos modèles issus de l’endosymbiose secondaire sont des microalgues marines (
Phaeodactylum tricornutum,
Microchloropsis gaditana), qui accumulent de l’huile en condition de stress, (notamment
M. gaditana) et pour lesquelles les outils modernes de manipulation génétique sont développés (CRISPR-CAS9). Les diatomées des glaces (
e.g. Fragilariopsis cylindrus) sont aussi étudiées.
Enfin, les glycérolipides membranaires et de réserve étant les premiers à répondre aux variations thermiques, et en particulier au froid, l’équipe porte une attention particulière aux organismes se développant dans la neige et la glace.
Nos questions scientifiques s’attèlent à :
1) caractériser des trafics et flux lipidiques entre organites,
2) décrypter le rôle et le contrôle de l’homéostasie lipidique à l’échelle de la membrane, de la cellule et de l’organisme,
3) reconstruire les voies de synthèses des lipides membranaires et
4) établir les relations entre la biosynthèse des lipides, l’expansion membranaire et l’architecture cellulaire.
5) comprendre le lien avec l’environnement abiotique des organismes, en particulier les milieux oligotrophiques pauvres en nutriments, et les variations thermiques, le froid, la neige, la glace.
Concernant le transfert des connaissances fondamentales vers les sciences appliquées, le contrôle de l’accumulation d’huile est étudié à travers le développement d’approches de génétique chimique et de biologie synthétique, en collaboration avec nos partenaires industriels.
Pour atteindre ses objectifs, l’équipe dispose des projets de recherche de chaque P.I., de l’accès au plateau technique d’analyse lipidomique, du développement des méthodes de microscopie électronique de pointe et de l’existence des deux partenariats industriels.
Quatre axes de recherches vont donc être développés dans les années à venir (Figure 2) :
- l’homéostasie des glycérolipides dans les endosymbiontes secondaires,
- le trafic lipidique chez les endosymbiontes primaires,
- le remodelage des glycérolipides chez les microalgues et les plantes en réponses aux changements de paramètres abiotiques, incluant ceux des environnements alpins,
- les réseaux de régulation de gènes sous-tendant la reprogrammation du métabolisme lipidique chez les microalgues marines.
