Le processus de photosynthèse mis en œuvre entre autres par les diatomées est une véritable prouesse biochimique car il permet la conversion d’une énergie électromagnétique – portée par des photons – en une énergie chimique directement utilisable par les cellules. Bien que la lumière soit une source d’énergie gratuite et abondante, elle est néanmoins intrinsèquement variable en intensité et en qualité, ce qui peut occasionner des dommages irréparables à l’appareil photosynthétique. Aussi, les organismes photosynthétiques ont développé des mécanismes d’acclimatation afin de dissiper l’énergie lumineuse en excès, à savoir le processus d'extinction non photochimique (
Non-Photochemical Quenching ou NPQ). En dépit de l’importance écologique des diatomées, les déterminants de la régulation du NPQ sont encore mal compris.
Les diatomées sont une famille remarquablement diversifiée de micro-organismes marins, capables de vivre aussi bien en eau douce et salée que sur la banquise. Leur forte présence dans les océans leur confère un rôle primordial dans le fonctionnement des écosystèmes marins, notamment en tant que puits de carbone et producteurs de dioxygène. Leur succès écologique est dû, entre autres, à l’exceptionnelle flexibilité de leur appareil photosynthétique qui leur permet de s’adapter à des changements de conditions lumineuses.
En utilisant une approche de spectroscopie, les chercheurs de notre laboratoire ont démontré qu’il existe chez les diatomées un couplage direct entre le NPQ et la composante ΔpH de la
force proton-motrice (PMF) générée par l’activité photosynthétique. Chez l’algue modèle
Phaeodactylum tricornutum, ils ont identifié l’existence d’un
antiport proton/potassium appelé KEA3. En s'avérant être un important régulateur de la PMF, cet antiport est indispensable pour que le NPQ s’établisse dans des conditions normales. En combinant génétique et photophysiologie, les chercheurs ont observé que l’antiport KEA3 est responsable de l’adaptation de la réponse NPQ aux conditions environnementales. En effet, KEA3 étant un échangeur d’ions, il est capable de convertir sans perte d’énergie la composante ΔpH de la PMF en ΔΨ. Ainsi, la PMF, et donc la production d’ATP, est maintenue tout en assurant une bonne protection de l’appareil photosynthétique grâce à la mise en place du NPQ.
Bien qu'elle soit apparentée sur le plan évolutif à la famille KEA1-3 rencontrée chez les plantes, la protéine KEA3 de diatomée contient un motif capable de lier un ion calcium. Les chercheurs montrent que ce domaine contrôle l'activité de KEA3 chez les diatomées, fournissant un lien possible entre la concentration intracellulaire de Ca
2+ et les réponses aux changements rapides (de l’ordre de quelques minutes) ou lents (par exemple journaliers) de l'environnement lumineux.
Dans l'ensemble, l’élucidation du circuit de régulation du NPQ chez les diatomées ainsi que celle du rôle de la protéine KEA3 peuvent contribuer à expliquer la prospérité de la famille des diatomées dans des environnements variés, où l'acclimatation à la lumière est souvent un déterminant majeur de la croissance et de la survie.
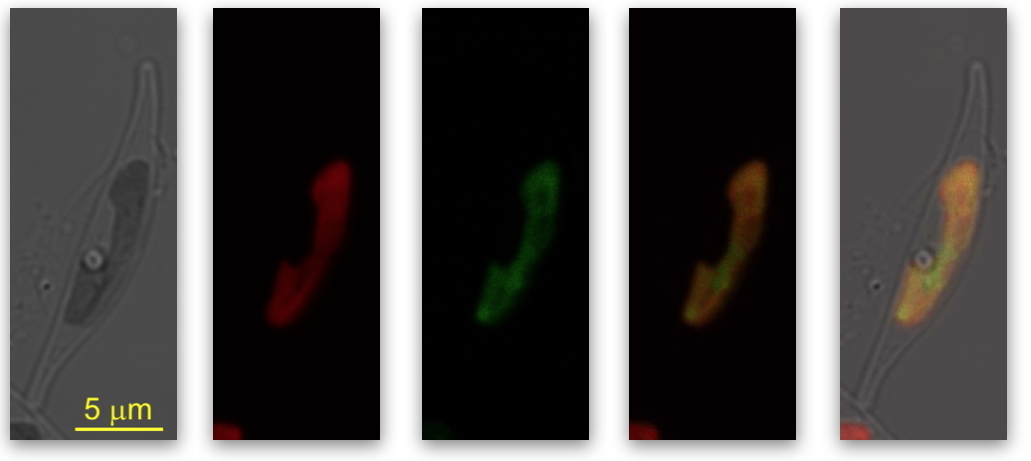
Localisation de la protéine KEA3 fusionnée dans
Phaeodactylum tricornutum.
1 - image en microscopie optique de la diatomée,
2 - auto-fluorescence de la chlorophylle,
3 - fluorescence de la protéine KEA3 fusionnée,
4 - fusion des images,
5 - fusion des images sur la microscopie optique.
La force proton-motrice correspond à un gradient de protons à travers une membrane biologique, de nature à la fois électrique (ΔΨ) et osmotique (ΔpH). Ce gradient peut être dissipé par le complexe transmembranaire ATP synthase dans les chloroplastes et les mitochondries afin de produire l’ATP, une molécule énergétique indispensable pour assurer le reste des processus biochimiques de la cellule.
Une protéine antiport est une protéine qui permet l’échange de deux molécules à travers la membrane cellulaire.