The photosynthetic membranes are so abundant (1 sqm of leaves contains a photosynthetic surface superior to that of a rugby stadium), that galactolipids, making the bulk of these membranes, are the most abundant polar lipids on Earth. These polar molecules consist of a hydrophilic headgroup and a hydrophobic tail consisting of 2 long carbon chains called fatty acids. Each cell membrane has its own glycerolipid composition: chloroplasts of plant cells and algae are characterized by the presence of a unique class of glycerolipids, the galactolipids. The de novo synthesis of fatty acids in a photosynthetic cell is initially carried out in the chloroplast stroma (whereas in animal cells it occurs in the cytoplasm). There are two main sites of assembly of membrane glycerolipids (directly generating biological membranes): the chloroplast envelope and the endoplasmic reticulum.
Galactolipids are synthesized in plant chloroplasts and harbor 2 fatty acids of 16 or 18 carbon atoms. In micro-algae and diatoms, galactolipids are rich in polyunsaturated acids with very long chains, including eicosapentaenoic acid, also known as EPA or 20:5, because it contains 20 carbons and 5 double bonds. If the chloroplast can theoretically synthesize fatty acids of 16 or 18 carbon atoms, it cannot produce longer species; the biosynthesis of 20:5 occurs therefore in the cytoplasm. Several enzymes generating 20:5 are known: elongases, which add 2 carbons to a fatty acid substrate, and desaturases, which add the double bonds. It was assumed that the initial substrate of the cytoplasmic enzymes was 18:2 subjected to one elongation step and three desaturations to reach 20:5. However, in microalgae such as Nannochloropsis, elongases are able to accept 16:0 as the initial substrate. What is the use of an elongation system that allows to pass from 16 to 18 carbon atoms in the cytoplasm? What is the role of these elongases?
To answer these questions, we have been interested in the family of 6 cytoplasmic elongases of Nannochloropsis (Δ0-ELO1-6) catalyzing the synthesis of 18:0 starting from 16:0. In a functional genetics study, the researchers showed that Δ0-ELO1, the most expressed of them, was the entry point of fatty acids to a multi-enzymatic system that leads to 20:5, directed then to the chloroplast where it is specifically used to produce a chloroplast galactolipid. From these data, the researchers obtained a mutant that did not express Δ0-ELO1 (Figure) and in which they observed the decrease in the content of this particular galactolipid, and by compensation, an increase in oil production overcoming the accumulation of 16:0.
This work not only provides answers to fundamental questions about the very complex synthesis of fatty acids in microalgae, but also highlights a way to genetically induce an accumulation of oil in Nannochloropsis. They are the subject of a patent application with the company Fermentalg.
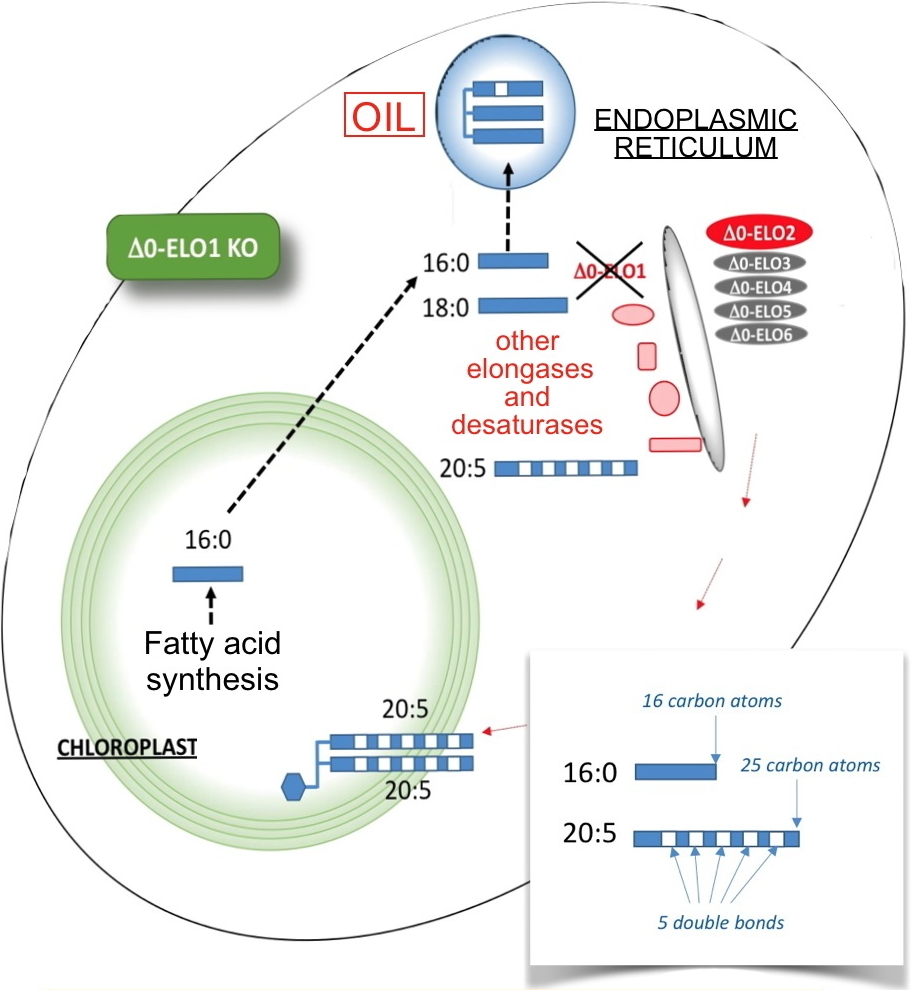
The chloroplastic 16:0 is normally exported to the cytosol where it is supported by elongases and desaturases to give 20:5 which is then redirected to the chloroplast. In this mutant that does not produce Δ0-ELO1, 16:0 accumulates and is in this case directed towards the production of oil.